|
page réalisée par Vincent ROBERT entomologiste médical directeur de recherche à l'I.R.D. chef du laboratoire de paludologie Institut Pasteur de Madagascar BP 1274, Antananarivo 101, Madagascar e-mail : robert@pasteur.mg mise à jour : 2 février 2001 |
|
le cycle de développement des anophèles le cycle trophogonique des anophèles la transmission homme-anophèle la transmission anophèle-homme les principales espèces vectrices en Afrique tropicale les principales espèces vectrices dans l'Océan Indien les principales espèces vectrices en Afrique du Nord la mesure du taux de transmission en zone d'endémie quelques exemples de taux de transmission les interactions entre transmission, morbidité et mortalité Une clé d'identification des anophèles : l'aile
- La transmission
par piqûre d'un moustique vecteur, appelée aussi transmission
vectorielle. C'est de loin le mode d'infection le plus important.
- La transmission
par transfusion sanguine avec du sang infecté.
- La transmission
trans-placentaire in utero, de la mère vers l'enfant.
Il existe environ 3300 espèces de moustiques appartenent à des genres différents : Aedes, Culex, Anopheles, etc. Parmi ceux-ci, seuls les anophèles sont potentiellement capables de transmettre le paludisme humain. On dénombre 422 espèces d'anophèles dans le monde, parmi lesquelles 68 ont été associées à la transmission des quatre espèces de paludisme humain. En Afrique et dans l'Océan Indien,une dizaine d'espèces seulment est concernée. Certaines de ces espèces peuvent également transmettre la filariose de Bancroft et des arboviroses. L'importance des anophèles est illustrée dans des zones comme la Nouvelle-Zélande, la Nouvelle-Calédonie, la Micronésie, la Polynésie, d'où les anophèles sont absents et qui, de ce fait, sont indemnes de paludisme. On remarque que le mot français anophèle est masculin (on dit un anophèle, même quand il s'agit d'une femelle). Ce mot anophèle correspond au latin Anopheles, désignant un genre de moustique, dont l'abréviation recommandée est An. Le cycle de développement des anophèles Le développement de toutes les espèces de moustiques est caractérisé par la succession de deux phases : la première est aquatique et recouvre la vie pré-imaginale, c'est-à-dire l'oeuf, les stades larvaires et la nymphe. La seconde est aérienne et concerne l'adulte, ou imago. Les anophèles femelles pondent sur l'eau des oeufs fécondés. Ces oeufs mesurent moins d'1 mm. Ils sont pondus isolémént, sont munis de flotteurs et restent en surface durant l'embryogénèse. De chaque oeuf éclôt une seule larve d'environ 1 mm. La larve aquatique des anophèles se nourrit en filtrant les débris organiques et les micro-organismes de l'eau. Il y a quatre stades larvaires, séparés par trois mues larvaires. La larve de stade 4 mesure 5 mm à 1 cm. La larve de quatrième stade effectue une mue particulière, la nymphose. Cette dernière libère une nymphe aquatique mobile, qui ne se nourrit pas. De profonds remaniements de la morphologie s'effectuent au cours du stade nymphal. Les organes propres au stade larvaire (appareil buccal filtreur-broyeur, système digestif de détritophage-filtreur) sont détruits ; les organes caractéristiques de l'adulte qui étaient présents à l'état d'ébauches dans la larve (ailes, pattes, appareil buccal piqueur-suceur, système digestif d'hématophage) apparaissent. De la nymphe émerge un adulte, mâle ou femelle, qui s'envole rapidement. C'est au cours de la phase aérienne, le plus souvent dans un essaim de mâles réunis au crépuscule, que s'effectue l'insémination de la femelle néonate. Les spermatozoïdes sont introduits dans la bourse copulatrice de la femelle, puis ils migrent dans une spermathèque, sorte d'annexe du système sexuel femelle, dans laquelle ils conservent leur pouvoir fécondant pendant plusieurs semaines, jusqu'à la mort de la femelle. Mâles et femelles se nourrissent de jus sucrés, nectars et autres exsudats végétaux. Le mâle est incapable de perforer la peau d'un vertébré. Seule la femelle est hématophage : un repas sanguin n'est pas indispensable à sa survie, mais il l'est à la maturation ovarienne, comprenant habituellement 150 oeufs chez Anopheles gambiae. La fécondation s'effectue dans le tractus génital de la femelle, lors de la ponte, des spermatozoïdes étant libérés à partir de la spermathèque. A la température de 25 °C, la durée de la phase pré-imaginale est d'une dizaine de jours pour Anopheles gambiae et d'une vingtaine paour Anopheles funestus. Cette phase s'allonge quand la température diminue et raccourcit quand elle augmente (cinq jours à 30 °C pour Anopheles gambiae). La longévité des anophèles femelles adultes est d'une vingtaine de jours, guère plus d'un mois dans les conditions habituelles. Elle est plus réduite pour le mâle. Il est généralement admis que la mortalité des anophèles femelles n'est pas constante, mais qu'elle augmente avec l'âge, tout particulièrement après une vingtaine de jours de vie imaginale pour Anopheles gambiae et Anopheles funestus. La dispersion moyenne des anophèles adultes en vol à partir des gîtes larvaires est de l'ordre de 1 km à 1,6 km, pouvant atteindre 3 km. La distance de 7 km est la limite maximale observée pour Anopheles gambiae. Dans les zones tropicales, la croissance de la plupart des espèces de moustiques est continue tout au long de l'année, avec des variations saisonnières de densité liées aux conditions climatiques. Pendant la saison sèche, notamment au Sahel et au Sahara, ces conditions deviennent franchement défavorables, entraînant une disparition apparente des populations anophéliennes. L'estivation, état physiologique de dormance permettant une survie prolongée de l'adulte en saison chaude dans l'attente du retour de la saison des pluies, a été démontrée uniquement pour Anopheles arabiensis au Soudan.
(source : J. Brunhes et Coll., Les anophèles de la région afro-tropicale, logiciel ORSTOM Ed., 1998) 1. L'oeuf : La forme de l'oeuf de l'anophèle évoque celle d'un cylindre incurvé aux extrêmités. Il mesure environ 0,5 mm de long et est muni de deux flotteurs latéraux remplis d'air. L'oeuf est protégé par plusieurs enveloppes particulières qui, contrairement à d'autres espèces de moustiques, ne lui permettent toutefois pas de résister à la sécheresse. 2. La larve : Elle se compose de trois parties : la tête, le thorax et l'abdomen. Chez Anopheles gambiae, elle mesure 1 mm de long au premier stade et 5 mm au quatrième. - La tête
comprend notamment les yeux, les antennes et les pièces buccales
qui entourent la bouche ventrale. Un cou membraneux sépare
la tête du thorax. Pour se nourrir, la larve effectue une rotation
de la tête de 180°, si bien que la bouche se retrouve juste
sous la surface de l'eau. Elle filte et ingère les particules
alimentaires amenées vers la bouche par le courant de surface
engendré par le battement des brosses buccales.
- Le thorax
est formé de trois segments non individualisés.
- L'abdomen,
cylindrique, se compose de neuf segments. Les sept premiers portent
notamment des plaques dorsales sclérifiées et des soies
palmées caractéristiques des anophèles. Ces soies
contribuent au maintien de la larve juste sous la surface de l'eau,
dans la position typique des anophèles, parralèle à
la surface de l'eau, face dorsale vers le haut. Le huitième
segment abdominal porte dorsalement deux orifices respiratoires, les
stigmates, s'ouvrant directement au niveau d'une plaque spiraculaire.
L'absence de siphon respiratoire différencie les anophèles
des autres moustiques. Les stigmates respiratoires sont ouverts en
surface pour autoriser le renouvellement d'air du système trachéen
et fermés par des valves lors de la plongée. La respiration
est aérienne. Le dernier segment abdominal porte l'anus.
3. La nymphe : En fin de quatrième stade larvaire, la cuticule se fend dorsalement et libère une nymphe, bien différente de la larve. Elle est composée de deux parties : le céphalothorax, résultant de la coalescence d'une tête non individualisée et d'un thorax globuleux, et l'abdomen. Le céphalothorax porte deux trompettes respiratoires, qui correspondent aux stigmates antérieurs du thorax de l'adulte. Ces trompettes à extrêmités hydrophobes traversent la surface de l'eau et assurent la respiration aérienne de la nymphe. L'abdomen comprend huit segments bien visibles, dont le huitième porte une paire de palettes natatoires. Lorsque l'abdomen se contracte brutalement, la nymphe effectue des mouvements saccadés. 4. L'adulte : Après quelques jours de vie nymphale, la cuticule du céphalothorax de la nymphe âgée se fracture dorsalement selon un plan sagittal. Les plans hydrophobes de la fracture écartent la surface de l'eau et libèrent successivement les différentes parties de l'adulte : thorax, tête, antennes, ailes, trompe, pattes, abdomen. L'adulte se retrouve à l'air libre, posé sur son exuvie nymphale. Il reste immobile, le temps que ses ailes se déploient et que la cuticule durcisse : après une heure, il est capable de s'envoler. L'émergence dure quelques minutes et représente une phase délicate dans la vie de l'insecte en raison d'une forte mortalité par noyade. L'adulte comprend trois parties bien distinctes : la tête, le thorax et l'abdomen. - La tête
porte deux gros yeux composés, une paire d'antennes de quinze
articles à soies nombreuses et longues chez le mâle,
rares et courtes chez la femelle. La trompe, ou proboscis, est formée
d'une pièce buccale particulière, le labium, formant
une gouttière souple entourant les autres pièces buccales
vulnérantes chez la femelle : l'hypopharynx, les deux mandibules
et les deux maxilles. Certaines de ces pièces buccales forment
deux canaux indépendants dans lesquels la circulation des fluides
se fait dans des sens bien définis et inverses l'un par rapport
à l'autre. Le plus gros est le canal alimentaire, délimité
par le labre replié en gouttière. Le moustique y aspire
le sang de l'hôte. Le plus fin est le canal salivaire, placé
à l'intérieur de l'hypopharynx : la salive est injectée
dans ce canal et circule depuis les glandes salivaires du moustique
jusqu'à la partie distale de l'hypopharynx, à l'extrêmité
la plus avancée du point de piqûre dans l'hôte.
Ces caractéristiques morphologiques permettent dans le même
temps à l'anophèle femelle, lors de la piqûre,
d'ingérer un volumineux repas sanguin et d'injecter avec sa
salive le stade infectant (sporozoïte) du Plasmodium.
Deux palpes maxillaires sont situés de part et d'autre de la
trompe. Chez la femelle, ils sont de la même taille que la trompe.
Chez le mâle, ils sont plus longs et la partie distale est renflée
en massue.
- Le thorax
est formé de trois segments portant chacun une paire de pattes.
Le premier est réduit. C'est sur le second, très développé
et renfermant de puissants muscles alaires, que s'insère une
paire d'ailes. Sur le troisième est placée une paire
d'haltères, ou balanciers, qui est l'homologue d'une paire
d'ailes postérieures atrophiées, et qui joue un rôle
dans l'équilibration du vol. Les six pattes, longues et graciles,
sont formées de neuf articles. Les ailes présentent
de nombreuses écailles claires et sombres : leur arrangement
sur le rebord costal est caractéristique des anophèles.
- L'abdomen
est constitué de dix segments, dont sept sont bien visibles.
Chaque segment est constitué d'une plaque chitineuse dorsale
et d'une plaque ventrale reliées par une membrane qui autorise
la dilatation de l'abdomen lors de la prise d'un repas de sang et
lors de l'élaboration de la ponte dans les ovaires. Les trois
derniers segments portent l'anus et les appendices génitaux,
ou génitalias.
L'ensemble trompe-tête-thorax-abdomen est dans le même alignement. Au repos, cet alignement détermine par rapport au support un angle aigu caractéristique des anophèles. Le cycle trophogonique des anophèles Le cycle trophogonique (ou gonotrophique) est la succession des phénomènes physiologiques qui se produisent chez le moustique entre deux repas de sang successifs. Trois phases sont distinguées : la recherche de l'hôte et sa piqûre par la femelle à jeun ; la digestion du sang qui se déroule dans le même temps que la maturation ovarienne ; la recherche du lieu de ponte de la femelle gravide. 1. La recherche de l'hôte et la piqûre : La femelle à jeun se met, de nuit, à la recherche d'un hôte à piquer. La localisation de ce dernier se fait à plusieurs niveaux. L'identification des facteurs d'attractivité et des mécanismes de localisation et de reconnaissance de l'hôte à piquer sont des domaines de recherche actuellement en plein essor : à longue distance, interviennent les stimuli olfactifs tels que des kairomones spécifiques d'espèces animales (ou humaine) ; à moyenne distance, d'autres attractifs interviennent, comme le gaz carbonique émis par la respiration ; à courte distance, la chaleur de l'hôte, l'humidité corporelle et divers facteurs visuels pourraient jouer un rôle important. Pour la piqûre proprement dite, la chaleur relative de l'hôte est un facteur primordial. L'espèce Anopheles gambiae est particulièrement attirée par les odeurs humaines de pieds, même à grande distance, et pique préférentiellement les pieds et les chevilles. L'action de la microfaune bactérienne de la peau a récemment été prouvée dans ces émanations. Contrairement à d'autres moustiques, Anopheles gambiae n'est pas attiré par les odeurs provenant de la respiration. La piqûre d'un anophèle survient pendant la nuit. En Afrique, quand on est piqué pendant la journée, on peut être assuré qu'il ne s'agit pas d'un anophèle. Il est possible d'être piqué par un anophèle infecté depuis le crépuscule jusqu'à l'aurore inclus. Ce risque est maximal au milieu de la nuit et au cours de sa seconde partie : en effet, les moustiques pares (i.e. les moustiques qui ont déjà pondu au moins une fois et qui sont les seuls à pouvoir être infectants) sont ordinairement occupés par la recherche du gîte de ponte et par la ponte au cours de la première partie de la nuit. Classiquement, la piqûre d'un anophèle s'effectue en une seule fois ; elle dure normalement deux à trois minutes, et permet la réplétion complète de l'estomac de la femelle. La plupart du temps, elle est réalisée sur un homme endormi. En ce sens, elle s'oppose à la piqûre d'autres espèces de moustiques, tels les Aedes, piquant pendant la journée et piquant de nombreuses fois la même personne ou des personnes différentes avant d'avoir ingéré un repas complet. Lors de la piqûre, le sang est ingéré par pompage après canulation directe d'un capillaire sanguin. Moins fréquemment, il peut aussi être aspiré dans une micro-hémorragie consécutive à la pénétration des pièces buccales vulnérantes. La salive est éjectée pendant toute la durée de la piqûre, depuis le début de la pénétration des pièces buccales jusqu'à la réplétion complète de l'estomac. La salive contient des anticoagulants, des substances inflammatoires, des enzymes digestives et des vasodilatateurs. Par définition, la tendance à piquer l'homme est l'anthropophilie, la tendance à piquer les animaux la zoophilie. 2. La digestion du sang et la maturation ovarienne : Dès la fin du repas commence la digestion du sang. La digestion d'un repas complet dure généralement une quarantaine d'heures. Le sang est d'abord concentré par élimination de l'eau : l'abdomen, distendu à l'extrême, qui paraît rouge par transparence à la fin du gorgement, devient brun-foncé et son volume diminue au fur et à mesure que la digestion progresse. Simultanément, les ovaires se développent. Les femelles pares nécessitent un seul repas de sang pour élaborer complètement une ponte. Les femelles nullipares ont parfois besoin de deux repas de sang pour achever le développement ovarien : on dit alors qu'elles présentent un stade prégravide. 3. La recherche du lieu de ponte et la ponte : Dès que la ponte est mature, la femelle se met en quête de gîtes favorables à la ponte. Comme pour la recherche de l'hôte à piquer, des stimuli olfactifs sont perçus à distance par la femelle et lui permettent de les localiser. A courte distance, des caractéristiques physio-chimiques de l'eau sont analysées. La ponte a généralement lieu peu après le crépuscule. Une fois cette dernière achevée débute un nouveau cycle. Pour les femelles nullipares (i.e. qui n'ont jamais pondu), le premier repas sanguin survient deux ou trois nuits après l'émergence. Pour les femelles pares (i.e. qui ont déjà pondu au moins une fois), la prise de repas de sang se fait généralement dans les heures qui suivent la ponte, au cours de la même nuit. La durée du cycle trophogonique est bien connue pour les vecteurs d'Afrique tropicale : elle varie entre deux et trois jours pouyr les femelles pares, suivant les espèces et les saisons. Pour les nullipares, il est plus long, jusqu'à cinq jours, du fait de la phase prégravide. La transmission homme-anophèles et le développement du paludisme dans le vecteur 1. Le cycle du parasite dans le vecteur : L'infection d'un anophèle femelle commence lors d'un repas sanguin pris sur un homme impaludé porteur de gamétocytes. Si d'autres stades parasitaires sont ingérés par le moustique, ils sont tués et digérés. Le stade gamétocyte est haploïde, comme tous les stades chez l'Homme. Il est rencontré dans le sang périphérique sous les formes mâle et femelle, ordinairement dans la proportion d'un mâle pour trois femelles. Les gamétocytes commencent un processus d'activation dans le bol alimentaire à l'intérieur de l'estomac du moustique. Ils acquièrent une forme sphérique et la membrane plasmique érythrocytaire se rompt. Le gamétocyte femelle activé est le macrogamète. Le gamétocyte mâle subit une exflagellation, processus de bourgeonnement de flagelles (jusqu'à huit, mais ordinairement de quatre à six), qui se détachent du corps du gamétocyte mâle activé et qui constituent autant de microgamètes. Toutes ces transformations interviennent une dizaine de minutes après la prise du repas sanguin. Le gamète mâle se déplace par ses propres mouvements, rencontre un gamète femelle puis pénètre dans ce dernier. Ce processus de fécondation aboutit à la formation d'un oeuf diploïde de forme sphérique. Une dizaine d'heures après le repas infestant, cet oeuf se transforme en un ookinète, via un stade intermédiaire dit "en virgule" (en anglais : retort) au cours duquel ce qui n'est pas encore l'ookinète bourgeonne à partir de ce qui n'est déjà plus l'oeuf. Cet ookinète est de forme allongée et présente les caractéristiques d'un stade invasif. Il se déplace par ses propres mouvements vers la périphérie du bol alimentaire, pénètre à travers l'enveloppe péritrophique (structure extracellulaire produite par l'estomac et enveloppant l'ensemble du bol alimentaire), puis traverse l'épithélium stomacal pour finalement s'immobiliser sous la lame basale de cet épithélium. Là, l'ookinète se dédifférencie en prenant une forme sphérique caractéristique d'un jeune stade oocyste. Cet oocyste s'accroît considérablement pendant une dizaine de jours en subissant de nombreuses multiplications nucléaires, dont la première est réductionnelle. A l'intérieur de cet oocyste se développent quelques 10 000 sporozoïtes qui, à maturité, sortent individuellement par effraction, pour se retrouver dans l'hémolymphe du moustique, puis dans toutes les parties du moustique. Ces trophozoïtes ont un tropisme positif pour les glandes salivaires dans lesquelles ils pénètrent, traversant de part en part l'épithélium des glandes salivaires et se retrouvant dans le canal salivaire, prêts à être injectés avec la salive lors du prochain repas de sang du moustique. Il semble acquis qu'un anophèle avec des sporozoïtes dans ses glandes salivaires reste infectant toute sa vie. 2. Les facteurs impliqués dans le succès du passage et du développement du parasite dans le vecteur sont nombreux et leur interaction complexe. Au moins six facteurs sont individualisés : - La température
figure au premier rang de ces facteurs : en dessous de 18 °C et
au dessus de 35 °C, le développement sporogonique de Plasmodium
falciparum est arrêté.
Aux températures de 20 °C, 24 °C et 30 °C, il est
respectivement de 20, 11 et 9 jours.
- La densité
gamétocytaire est également un facteur ayant une influence
évidente et positive sur l'infection des vecteurs.
- L'état
immunitaire de l'homme porteur de gamétocytes : des anticorps
dirigés contre le parasite aux stades précoces du développement
sporogonique peuvent bloquer, partiellement ou même totalement,
la transmission du parasite vers le moustique, à l'intérieur
de ce dernier.
- Le statut
drépanocytaire du porteur de gamétocytes : l'hémoglobine
AS favorise le passage du parasite vers le moustique.
- L'âge
du gamétocyte : un âge de 3 à 5 jours, après
l'observation des premières exflagellations, semble optimum.
- Le sex ratio
des gamétocytes : une relativement forte proportion de mâles
est favorable à l'infection des moustiques.
3. Le parasite est-il pathogène pour le moustique ? Dans l'état actuel de nos connaissances, on peut répondre par la négative à cette question, au moins pour Anopheles gambiae, qui semble parfaitement tolérer Plasmodium falciparum. Le développement sporogonique du parasite et son maintien au stade sporozoïtes dans les glandes salivaires ne semble pas avoir de répercussion sur la survie du moustique infecté. Un faible effet tératogène n'est cependant pas exclu, mais il serait sans implication épidémiologique majeure. Un tel effet expliquerait par exemple pourquoi on n'observe que très rarement dans la nature des anophèles avec un grand nombre d'oocystes, alors qu'à l'insectarium cette observation est fréquente. Par ailleurs, on a l'assurance que certaines espèces plasmodiales ne parasitant pas l'homme sont effectivement pathogènes pour leurs vecteurs. La transmission anophèles-homme La piqûre de l'anophèle porteur de sporozoïtes dans ses glandes salivaires est un des rares moments de rencontre entre les trois acteurs du paludisme : l'homme, le vecteur et le parasite. C'est à cet instant que s'effectue l'infection de l'homme. La mobilité des sporozoïtes, quoique avérée, est trop réduite pour jouer un rôle dans le processus de passage du vecteur à l'hôte. Le transfert des sporozoïtes des glandes salivaires à l'hôte humain est considéré comme passif, totalement dépendant de l'accès des sporozoïtes à la salive et du flot de cette dernière lors de la piqûre. Le nombre de sporozoïtes présents dans les glandes salivaires d'un anophèle infecté peut être très variable, d'une dizaine à plusieurs centaines de milliers. En général, la moitié des anophèles héberge moins de 1000 sporozoïtes dans les glandes salivaires. Cependant, le nombre de sporozoïtes effectivement injectés dans l'hôte lors de la piqûre est étonnamment faible, de l'ordre d'une dizaine, pouvant exceptionnellement atteindre plusieurs centaines. Ce nombre de sporozoïtes injectés représente environ 1 % du nombre de sporozoïtes contenus dans les glandes salivaires. Les sporozoïtes injectés dans la peau pendant la pénétration des pièces buccales (i.e. avant le gorgement de sang) semblent les seuls à pouvoir parvenir dans le foie de l'hôte. Les sporozoïtes injectés dans le vaisseau sanguin pendant le gorgement sont sans avenir pour le cycle parasitaire car ils sont aussitôt réingérés par le moustique dans son repas de sang. C'est pourquoi des sporozoïtes sont normalement retrouvés dans le contenu stomacal d'un anophèle infectieux et fraîchement gorgé. Le succès d'une seule piqûre pour infecter un hôte non immun n'est jamais absolu : quoique encore imparfaitement évalué, il avoisinne probablement une chance sur deux. Le comportement de piqûre de l'anophèle avec des sporozoïtes dans les glandes salivaires semble modifié, selon les espèces de vecteurs et peut-être aussi selon l'espèce plasmodiale. On observe pour ces anophèles une augmentation du temps de pénétration des pièces buccales et/ou une augmentation du nombre moyen d'hôtes piqués pour atteindre une réplétion complète. Ces modifications du comportement des anophèles semblent attribuables à la présence du parasite dans les glandes salivaires : elles vont clairement dans un sens favorisant la transmission du parasite. Les principales espèces d'anophèles vectrices en Afrique tropicale Le complexe Anopheles gambiae et l'espèce Anopheles funestus assurent la majeure partie de la transmission en Afrique tropicale. Anopheles nili et Anopheles moucheti y jouent un rôle important dans des régions limitées. D'autres espèces peuvent avoir un rôle vecteur secondaire, localement, dans les régions où elles abondent, bien que l'essentiel de la transmission reste assuré par les espèces pré-citées. C'est à cause de la grande capacité vectrice de ces anophèles que l'endémie palustre est si forte en Afrique. 1. Le complexe Anopheles gambiae : Les différentes espèces du complexe Anopheles gambiae (ou Anopheles gambiae sensu lato) ont en commun une morphologie très semblable, voire identique : on parle d'espèces jumelles. Les déterminations à l'intérieur du complexe se fondent essentiellement sur trois critères : Des critères
de reproduction en insectarium, avec des espèces de référence.
Les croisements entre espèces différentes donnent naissance
à des mâles hybrides vivants mais stériles. Au
contraire, les femelles, même hybrides, sont parfaitement fécondes.
Des critères
cyrogénétiques utilisant la séquence des bandes
claires et sombres de chromosomes particuliers, les chromosomes géants
ou polyptènes. Ces chromosomes sont observés soit dans
les glandes salivaires des larves, soit dans les cellules nourricières
des ovaires de la femelle semi-gravide.
Des critères
moléculaires, tels que le polymorphisme de longueur des séquences
de DNA microsatellites, révélé par la technique
PCR.
On dénombre sept espèces, toutes africaines, dans ce complexe : - Anopheles
gambiae sensu stricto
- Anopheles
arabiensis
- Anopheles
quadriannulatus espèce A
- Anopheles
quadriannulatus espèce B
- Anopheles
bwambae
- Anopheles
melas
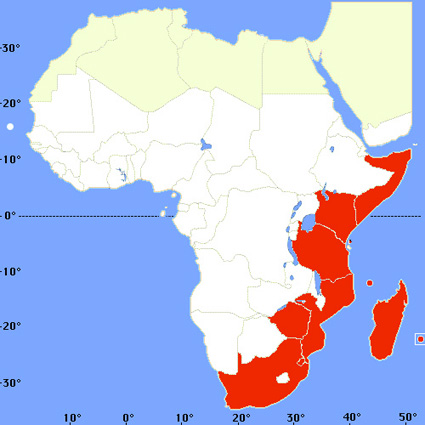
- Anopheles merus Anopheles gambiae sensu stricto et Anopheles arabiensis présentent les plus vastes aires de distribution. Distribution d'Anopheles gambiae et d'Anopheles arabiensis
(source : J. Brunhes et Coll., Les anophèles de la région afro-tropicale, logiciel ORSTOM Ed., 1998) Ces deux espèces vivent dans la quasi totalité de l'Afrique subsaharienne non méridionale. Elles ne dépassent pas 1000 mètres d'altitude. Anopheles arabiensis est absent de la forêt dense. Ces deux espèces ont une grande amplitude écologique. Classiquement, Anopheles gambiae domine en zones de forêt et de savane humide, tandis qu'Anopheles arabiensis est avantagé dans les savanes plus sèches, les steppes et parfois les villes, notamment dans le sud du Nigeria. Les gîtes larvaires sont des collections d'eau peu profondes et ensoleillées : empreintes de pas, flaques, petites mares, marécages aménagés, rizières, flaques résiduelles des cours d'eau en décrue. Les larves d'Anopheles gambiae et d'Anopheles arabiensis se rencontrent aussi dans d'autres types de gîtes, mais de manière inhabituelle. Elles ne se rencontrent en principe pas dans les eaux fortement ombragées, à courant rapide, alcalines ou polluées. La densité des populations varie selon la saison en fonction de la pluviosité. Ainsi, en zone de savane à une seule saison des pluies, elle s'accroît rapidement dès les premières pluies et la densité maximale est atteinte dans la seconde moitié de la saison pluvieuse. Elle décroît ensuite. Les femelles piquent de préférence au niveau du sol, à l'intérieur des maisons (on parle d'endophagie de piqûre). Mais l'exophagie n'est pas rare, en particulier chez Anopheles arabiensis. Les deux espèces sont nettement anthropophiles, sauf si l'abondance de bétail à proximité (boeufs surtout, mais aussi chèvres et moutons) favorise la zoophilie. Classiquement, l'activité de piqûre augmente au cours de la première moitié de la nuit, atteint sa valeur maximale entre minuit et quatre heures, puis diminue en persistant jusqu'à l'aube. Anopheles arabiensis et surtout Anopheles gambiae aiment à séjourner à l'intérieur des cases (on parle d'endophilie). L'endophilie-exophilie est variable selon la saison, les préférences trophiques, l'habitat, etc. D'une manière générale, l'endophilie est favorisée par la saison froide, l'anthropophilie et les zones arides sans lieux favorables à l'extérieur pour leur repos diurne. La dispersion active est de l'ordre de 2 km. Le cycle trophogonique dure trois à cinq jours chez les femelles nullipares et de deux à trois jours chez les pares. Parmi les autres espèces du complexe, seules Anopheles bwambae, Anopheles melas et Anopheles merus jouent un rôle, secondaire, dans la transmission du paludisme. On rencontre Anopheles bwambae dans une forêt d'Ouganda, Anopheles melas sur le littoral atlantique, Anopheles merus sur le littoral de l'Océan Indien. Ces trois espèces utilisent des gîtes larvaires à forte teneur en sel : les larves d'Anopheles bwambae se développent dans des sources d'eau minérale, celles d'Anopheles melas et d'Anopheles dirus dans de l'eau saumâtre (lagunes). Distribution d'Anopheles melas
(source : J. Brunhes et Coll., Les anophèles de la région afro-tropicale, logiciel ORSTOM Ed., 1998)
Distribution d'Anopheles merus
(source : J. Brunhes et Coll., Les anophèles de la région afro-tropicale, logiciel ORSTOM Ed., 1998) 2. Le groupe Anopheles funestus : L'espèce Anopheles funestus est répandue dans toute l'Afrique subsaharienne. Elle est abondante dans les savanes ouvertes. Elle atteint 2000 mètres en montagne, où sa densité est souvent supérieure à celle d'Anopheles gambiae s.l. Ses gîtes larvaires sont typiquement des gîtes d'eaux profondes, claires, permanentes ou sub-permanentes, ombragées par la végétation (herbes, végétation flottante). Ce sont des mares, des marécages, des bordures de lacs et de cours d'eau. Les rizières sont utilisées par ce moustique en Afrique de l'Est et à Madagascar, mais pas en Afrique de l'Ouest. La densité de la population imaginale varie avec la pluviosité, mais avec un certain retard par rapport à Anopheles gambiae s.l.. Ce retard est dû à deux facteurs : d'une part, la mise en condition des gîtes d'eaux profondes, d'autre part le développement pré-imaginal relativement lent. En zone de savane, la densité des populations d'adultes augmente bien après le début des pluies et atteint son maximum au début de la saison sèche, au moment où les populations d'Anopheles gambiae s.l. ont déjà sensiblement diminué. En zone de forêt, plus régulièrement et davantage arrosée, les densités sont plus stables et généralement plus faibles. Cette espèce, comme Anopheles gambiae et Anopheles arabiensis, préfère de beaucoup piquer l'homme, mais un certain opportunisme tend à augmenter la zoophilie lorsqu'il y a abondance de bétail sensible : on parle de déviation trophique. L'endophagie est très marquée. En général, l'activité de piqûre débute au crépuscule, croît et atteint son maximum dans les quatre dernières heures de la nuit. En saison froide, ce pic se situe au début de la nuit, alors que la température n'a que peu diminué. Les femelles se reposent habituellement à l'intérieur des habitations. Cependant, les femelles nullipares sont plus exophiles. Le cycle trophogonique dure en général quatre à cinq jours chez les femelles nullipares et deux à trois jours chez les pares. La distance de vol entre lieux de piqûres et de pontes est de l'ordre de 2 km, mais elle peut atteindre 7 km. Distribution d'Anopheles funestus
(source : J. Brunhes et Coll., Les anophèles de la région afro-tropicale, logiciel ORSTOM Ed., 1998) Une autre espèce appartenant à ce groupe, Anopheles rivulorum, quoique peu anthropohile, s'est avérée vectrice de paludisme en Tanzanie. 3. Anopheles nili : Anopheles nili est très largement répandu dans toute l'Afrique et est localisé aux environs des cours d'eau permanents ou semi-permanents dans lesquels se développent ses larves. En zone de savane, il est abondant au cours de la seconde moitié des pluies. Au bord du fleuve Congo, sa densité maximale est observée pendant la saison sèche, les larves étant emportées par le courant en saison des pluies. Au bord du fleuve Sanaga, dans le sud du Cameroun, sa densité est au contraire proportionnelle au débit du fleuve. Cette espèce pique volontiers l'Homme. L'activité de piqûre se manifeste assez rapidement après la tombée de la nuit et présente un pic plus précoce que ceux d'Anopheles gambiae et d'Anopheles funestus. Anopheles nili est nettement plus exophage et exophile que ces deux derniers. Distribution d'Anopheles nili
(source : J. Brunhes et Coll., Les anophèles de la région afro-tropicale, logiciel ORSTOM Ed., 1998) 4. Anopheles moucheti : Anopheles moucheti est limité au bloc forestier d'Afrique Centrale et aux galeries forestières qui le prolongent au nord et surtout au sud. Il est connu au sud Cameroun, au Gabon, au sud de la République Centrafricaine et au Congo. Les larves se rencontrent sur les bords des cours d'eau relativement larges, dans des eaux apparemment immobiles où la végétation abonde. Au voisinage des villages, les femelles adultes piquent aussi bien à l'intérieur qu'à l'extérieur des maisons, pendant toute la nuit. L'exophilie est souvent très marquée. Distribution d'Anopheles moucheti
(source : J. Brunhes et Coll., Les anophèles de la région afro-tropicale, logiciel ORSTOM Ed., 1998) Les principales espèces d'anophèles vectrices dans l'Océan Indien - Anopheles gambiae est observé sur le pourtour de Madagascar et dans l'archipel des Comores. - Anopheles arabiensis est observé sur les Hautes-Terres de Madagascar où il utilise les rizières, ainsi que dans le sud et l'ouest de Madagascar, l'archipel des Comores, Mayotte, La Réunion et Maurice. - Anopheles merus est un vecteur secondaire, présent dans le sud de Madagascar. - Anopheles funestus est partout présent à Madagascar. Il utilise les rizières comme gîtes larvaires. C'est le vecteur responsable des épidémies d'altitude. Il est présent aussi à Mayotte, Mohéli et Anjouan. - Anopheles mascarensis est un anophèle endémique, présent dans tout le sous-continent malgache. C'est un vecteur avéré sur la côte dans l'île de Sainte-Marie et il est même le vecteur principal dans la région de Fort-Dauphin. Etant très zoophile en altitude, il n'est alors plus vecteur. Il n'y a pas de vecteur du paludisme à Rodrigues, dans l'archipel des Chagos, dans l'archipel des Seychelles ni à Saint-Brandon. Les principales espèces d'anophèles vectrices en Afrique du Nord - Anopheles pharoensis : c'est un anophèle relativement grand, qui est très abondant dans le delta du Nil. Sa dispersion moyenne à partir des gîtes larvaires est considérable, de l'ordre de 6 km. De manière occasionnelle, en utilisant un vent du nord-ouest, ses déplacements ont été documentés sur plus de 100 km, une fois même sur 280 km, piquant en grand nombre des militaires en plein désert. Il est vecteur en Egypte. il est également présent en Afrique tropicale où son rôle vecteur est très faible et même totalement négligeable là où d'autres vecteurs majeurs sont présents. - Anopheles labranchiae est vecteur au Maghreb. - Anopheles sergentii est vecteur dans les zones irriguées de Lybie et d'Egypte. La mesure du taux de transmission en zone d'endémie Le taux d'inoculation entomologique (entomological inoculation rate, EIR) est considéré comme le paramètre entomologique reflétant le mieux la transmission du paludisme. Par définition, il est le résultat du produit entre le taux de piqûres (une estimation de la densité de vecteurs par homme) et l'indice sporozoïtique (une estimation de la proportion de vecteurs avec des parasites à un stade infectieux dans les glandes salivaires). Le taux d'inoculation entomologique est exprimé en nombre de piqûres d'anophèles infectés par homme et par unité de temps (une nuit, un mois, une année). La mesure du taux d'inoculation entomologique est considérée avec une certaine confiance dans les zones endémiques. Mais il n'en va pas de même dans les zones où la densité anophélienne et/ou l'indice sporozoïtique sont très faibles. on doit alors considérer le grand intervalle de confiance bornant la valeur moyenne du taux d'inoculation. Parfois, ce taux ne peut être calculé et l'estimation de la transmission se fait alors sur de toutes autres bases, parasitologiques, cliniques ou immunologiques. Dans les zones où la transmission est moyenne, le taux annuel d'inoculation entomologique n'est pas suffisant pour caractériser la transmission : la dynamique saisonnière de la transmission doit aussi être prise en compte. Considérons à titre d'exemple théorique deux zones où le taux d'incoculation entomologique est de 12 piqûres d'anophèles infectés par homme (PAIH) par an. Dans la première zone, 1 PAIH est régulièrement observée pendant tous les mois de l'année, tandis que dans la seconde, on relève 12 PAIH pendant un mois et aucune les onze autres mois : dans ces deux zones, l'épidémiologie du paludisme est radicalement différente. La transmission détermine l'intimité du contact entre l'Homme et son parasite, ce qui permet plus ou moins rapidement l'instauration à terme d'un équilibre de prémunition. Au contraire, dans les zones de très forte ou de très faible transmission, la dynamique saisonnière de la transmission est nettement moins importante : une transmission très élevée est forcément permanente ou sub-permanente, une transmission très faible forcément occasionnelle. Il est aussi important de noter que le taux d'inoculation entomologique ne peut être une mesure exacte de la transmission. En effet, beaucoup de piqûres d'anophèles infectés par des sporozoïtes dans les glandes salivaires échouent pour infecter un homme totalement dépourvu d'immunité anti-palustre. Des transmissions expérimentales ont montré qu'en moyenne un volontaire sur deux dévelopait une parasitémie après une unique piqûre d'anophèle infecté. Quelques exemples de taux de transmission Le taux d'inoculation entomologique est extrêmement variable. Les valeurs extrêmes observées sont 0, dans les zones indemnes de transmission, et 1000 dans certaines zones rurales. 1. En forêt dense équatoriale, non dégradée (de laquelle l'Homme est pratiquement absent), la transmission du paludisme est très faible ou nulle. Ceci s'explique par deux raisons : les vecteurs majeurs du paludisme sont très peu nombreux, la forêt dense leur étant manifestement défavorable ; en outre, l'Homme, à l'exception des Pygmées, est pratiquement absent de cette zone. Il n'y a donc pas de réservoir de parasites. La transmission est faible, de l'ordre d'une dizaine de PAIH (piqûres d'anophèles infectés par homme) par an. 2. En forêt dégradée, le long d'un fleuve ou d'une route, la situation est radicalement différente. La déforestation, naturelle ou provoquée par l'Homme, a un fort impact sur la transmission du paludisme. Les rayons du soleil pénètrent jusque dans les gîtes larvaires. Dès lors, ces gîtes deviennent favorables au développement d'Anopheles gambiae, qui peut devenir très abondant. Les conditions de survie (température, hygrométrie) étant ordinairement favorables à la survie de l'adulte, la transmission est très élevée, fréquemment de l'ordre de plusieurs centaines de PAIH par an. La transmission est permanente tout au long de l'année. 3. En savane, au contraire, les variations saisonnières sont marquées. La transmission survient uniquement au cours de la saison pluvieuse et au début de la saison sèche. Elle peut être considérable, variant entre plusieurs centaines et plusieurs dizaines de PAIH selon les localités. On observe fréquemment une succession des rôles vecteurs entre Anopheles gambiae et/ou Anopheles arabiensis en milieu de saison des pluies, puis Anopheles funestus en début de saison sèche. 4. Au Sahel, les conditions climatiques sont ordinairement trop rudes pour permettre une transmission au delà de deux mois en saison des pluies. Localement, notamment aux alentours des mares, la période de transmission peut être rallongée. La transmission est de l'ordre d'une à quelques dizaines de PAIH par an. 5. Au Sahara, la transmission du paludisme est un évènement anormal, d'ailleurs relativement peu documenté. Elle n'intervient que sur le mode épidémique dans les localités les plus favorables, i.e. les oasis, éventuellement à la faveur de l'augmentation du réservoir de parasites consécutif à l'apport d'immigrants impaludés. 6. En zone irriguée, ordinairement liée à des aménagements hydro-agricoles (par exemple les rizières dans les zones de savane, les modifications de l'environnement entraînent des répercussions très variables. Dans tous les cas, il y a augmentation de la densité des vecteurs du paludisme. Mais il n'y a pas toujours augmentation de la transmission : cela dépend du niveau de base de la transmission avant la mise en place de ces aménagements. Lorsque ce niveau de base est faible, il n'est pas rare de voir augmenter la transmission du paludisme. Au contraire, lorsque ce niveau de base est déjà important, les conséquences de l'aménagement sur la transmission sont faibles ou nulles. 7. En ville, la transmission du paludisme est une réalité parfaitement perceptible. Ellee est ordinairement de l'ordre de 0,1 à 3 PAIH par an. Ces taux sont les plus faibles de ceux observés en Afrique tropicale. C'est l'urbanisation par elle-même qui est responsable de ce bouleversement de l'épidémiologie du paludisme. Un gradient de transmission est ordinairement observé depuis les quartiers récents les plus périphériques, où la transmission est à peine plus faible que dans la zone rurale avoisinante, vers les quartiers anciens et centraux, où la transmission est beaucoup plus faible. Il n'est maintenant plus exceptionnel de rencontrer des hommes adultes ayant constamment vécu en milieu urbain et ayant échappé, par chance, à toute impaludation. Sans expérience palustre, ils sont dans un état immunologique proche de celui d'un voyageur venant d'un pays indemne de paludisme. Il est prévisible que cette situation naissante se développe avec l'urbanisation croissante, aussi bien en valeur absolue qu'en pourcentage de la population concernée, et que ces nouvelles situations potentiellement épidémiques devront être considérées à moyen et à long terme. 8. Il est intéressant de noter que c'est la valeur du taux de transmission qui détermine le contexte épidémiologique. En effet, par définition, le paludisme est dit ENDEMIQUE quand sa transmission est plus ou moins constante et perceptible pendant plusieurs années consécutives. Au contraire, il est dit EPIDEMIQUE lorsque sa transmission est inhabituelle, soit par son intensité au cours d'une période exceptionnelle, soit par sa localisation dans une zone où elle ne se déroule habituellement pas. Les interactions entre transmission, morbidité et mortalité Cette question est complexe : il n'existe pas une relation rectiligne entre la transmission et ses conséquences sur la santé. Aborder les relations entre la transmission, la morbidité et la mortalité revient d'emblée à distinguer : - Les zones d'endémie moyenne ou forte, dans lesquelles les taux de transmission sont toujours supérieurs à une PAIH (piqûre d'anophèle infecté par homme) par an. - Les zones de faible endémie, dans lesquelles la transmission est plus faible que dans les zones précédentes, avec parfois des variations inter-annuelles importantes. - Les zones épidémiques, dans lesquelles la transmission est un phénomène anormal auquel on a des chances significatives d'échapper au cours d'une vie humaine. Dans les zones de forte transmission, une seule piqûre infectée entraîne exceptionnellement un accès palustre. Cela est dû à l'acquisition progressive d'une prémunition, état immunitaire particulier qui se traduit par une protection relativement efficace par rapport à la maladie, mais insuffisante pour éliminer tous les parasites. Cette prémunition ne peut se construire et s'entretenir qu'à la faveur de réinfections multiples, presque permanentes, entre l'Homme et son parasite. Dans ces zones de forte transmission, elle s'établit dès le plus jeune âge, en quelques années, si bien qu'on estime qu'à l'âge de 5 ans, le risque de décéder du paludisqme est devenu très faible. On comprend alors pourquoi, dans ce contexte de transmission intense et permanente, l'essentiel de la morbidité et la quasi totalité de la mortalité sont concentrés chez les jeunes enfants. dans les zones de faible endémie et dans les zones épidémiques, la situation est bien différente. une unique piqûre infectée entraîne fréquemment un accès palustre. Le contact entre l'Homme et son parasite est trop peu fréquent pour que se construise l'état immunitaire de prémunition. Toutes les classes d'âge sont concernées par la maladie palustre et le risque de décès. dans tous les cas, il n'y a pas de taux de transmission qui soit acceptable en terme de santé publique. Quelque soit le taux de transmission, les conséquences sont toujours dramatiques sur tout ou fraction de la population humaine. Une clé d'identification des anophèles : l'aile Anopheles gambiae
Anopheles funestus
Anopheles moucheti
Anopheles nili
(source : J. Brunhes et Coll., Les anophèles de la région afro-tropicale, logiciel ORSTOM Ed., 1998) La transmission du paludisme est assurée par des moustiques du genre Anopheles. Le cycle de développement des anophèles est complexe : il comprend une première phase aquatique et une seconde, aérienne. A l'émergence, un anophèle adulte n'est jamais infecté par le paludisme. La partie du cycle parasitaire qui se déroule dans l'anophèle est, elle aussi, très complexe. Les transformations et les multiplications du parasite durent de 10 à 14 jours. Le passage du parasite de l'Homme au vecteur, puis du vecteur à l'Homme, se fait à l'occasion de piqûres. Le moustique mâle n'étant pas hématophage, seule la femelle est impliquée dans la transmission. La bio-écologie des vecteurs majeurs est très particulière : l'anthropophilie et l'endophagie de piqûre sont élevées et garantissent une forte interaction Homme-moustique. Il en résulte que les vecteurs du paludisme en Afrique tropicale ont une remarquable aptitude à transmettre : les taux de transmission du paludisme en Afrique intertropicale sont les plus élevés du monde. C'est principalement pour cette raison que l'endémie palustre est si élevée dans cette zone. Il n'y a pas de transmission du paludisme qui soit acceptable en terme de santé publique. La seule façon absolue de prévenir le paludisme est de ne pas recevoir de piqûres d'anophèles infectés. En Afrique tropicale, il n'existe probablement aucune localité où l'on puisse être sûr à 100 % de l'absence de transmission. Tout voyageur, dès qu'il passe une fraction de nuit dans cette partie du monde, même en simple escale sans descendre de l'avion, prend le risque d'être piqué par un anophèle infecté. Des comportements et des pratiques préventives peuvent significativement réduire ce risque (cf : chapitre sur la protection individuelle et la lutte anti-vectorielle), mais jamais l'annuler complètement. BEIER J-C., VENDERBERG J-P. - Sporogonic development in the mosquito. In : Malaria : parasite biology, pathogenesis, and protection ; Irwin W. Sherman Ed., ASM Press, Washington 1998 : 49-61. GILLIES M.T., COETZEE M. - A supplement to the Anophelinae of Africa South of the Sahara (Afrotropical region). Publ. South Afr. Inst. Med. Res. ; Johannesburg 1987 ; n°55, 143 p. GILLIES M.T., DE MEILLON B. - The Anophelinae of Africa South of the Sahara (Ethiopian zoogeographical region).Publ. South Afr. Inst. Med. Res. ; Johannesburg 1968 ; n°54, 2ème édition, 343 p. KETTLE D.S. - Medical and veterinary entomology. CAB International, Wallingford UK 1995, 2ème édition, 725 p. MOUCHET J., CARNEVALE P., COOSEMANS M. et Coll. - Typologie du paludisme en Afrique. Cahiers Sante 1993 ; 3 : 220-238. ROBERT V. - Les relations anophèles-plasmodium : conséquences pour la transmission du paludisme. Malaria and Infectious Diseases 1999 ; 9bis : 22-35. ROBERT V., BREY P.T. - Biting physiology of Anopheles affecting Plasmodium transmission. Res. Rev. Parasitol. 1999 ; sous presse. RHODAIN F., PEREZ C. - Précis d'entomologie médicale et vétérinaire. Notions d'épidémiologie des maladies à vecteur. Maloine Ed., Paris 1985, 458 p. TAKKEN W., KNOLS B.G. - Odor-mediated behavior of afrotropical malaria mosquitoes. Annual review of Entomology 1999 ; 44 : 131-157. ZAHAR A.R. - Vector bionomics in the epidemiology and control of malaria. Part 1. The WHO African region and the southern WHO eastern mediterranean region. WHO-VBC/84.3, 1984, 109 p. |